
Оваријум
Последње прегледано: 23.04.2024

Сви иЛиве садржаји су медицински прегледани или проверени како би се осигурала што већа тачност.
Имамо стриктне смјернице за набавку и само линкамо на угледне медијске странице, академске истраживачке институције и, кад год је то могуће, медицински прегледане студије. Имајте на уму да су бројеви у заградама ([1], [2], итд.) Везе које се могу кликнути на ове студије.
Ако сматрате да је било који од наших садржаја нетачан, застарио или на неки други начин упитан, одаберите га и притисните Цтрл + Ентер.
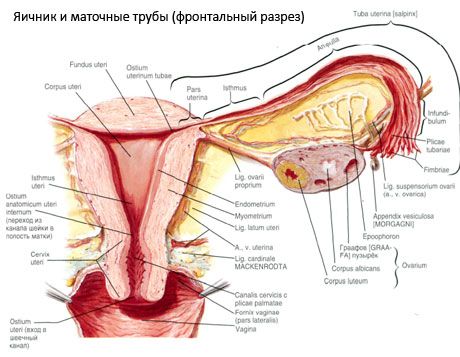
Оваријум (оваријум, грчки оофхорон) - упарени орган, женска генитална жлезда, смештена у шупљини мале карлице иза широког лигамента материце. Оварије развијају и сазревају женске полне ћелије (овуле), као и женске полне хормоне који улазе у крв и лимф. Оваријум има овалан облик, донекле равнициран у предњем и задњем правцу. Боја јајника је ружичаста. На површини јајника жене која рађа, видљиве су депресије и ожиљци - трагови овулације и трансформације жутих тела. Маса јајника износи 5-8 г. Димензије јајника су дуге 2,5-5,5 цм, широке 1,5-3,0 цм и дебљине 2 цм. Јаја има две слободне површине: фациал медиалис ) окренут према карличне шупљине, делимично покривена јајовод, и бочној површини (фациес латералис), уз карличног бочног зида, благи продубљивања - јајника фосса. Ова фовеа лежи у углу између перитонеалних спољашњих илиакова посуда на врху и материце и оклузалних артерија на дну. Иза јајника, уретер одговарајуће стране пролази од врха до дна.
Јајника површина креће конвексног слободном (позади) ЕДГЕ (марго Либер), испред - у мезентеричној региону (марго месоварицус), помоћу кратког клипа износа перитонеума (мезентеријуму овари) за полеђину листа широких лигамента материце. На челу тела је Флутед одмор - јајника капија (хилум оварии), кроз који јајник укључују артерије и нерве налази вене и лимфатике. Ми такође изолован јајника два краја: заобљени горњи цевасти завршетак (ектремитас тубариа), окренут јајовод, а доњи крај мајке (ектремитас утенна), заједно са материце сопствене Бунцх јајника (лиг оварии сопствене.). Овај скуп у округлом кабла око 6 мм од мајке је крај на бочном углу јајника материце, која се налази између два листа широке лигамент. Би лигамента апарата јајнику такође односи лигамента подвесхиваиасцхаиа јајник (лиг.суспенсориум оварии), што представља износа перитонеума протеже од горњег зида карлице у јајник и јајника садрже судова и фиброзних сноповима влакана. Јајник фикед схорт мезентеријум (месовариум), која представља дупликатури перитонеум, протеже од задње летка широког лигамента материце до јајника мезентеријалне ивице. Сами јајника нису покривени перитонеумом. До краја цеви је приложен јајника веома велике јајника Фимбриа на јајовод. Топографија јајника зависи од положаја материце, вредност (у трудноћи). Оварије се односе на веома мобилне органе карличне шупљине.
 [
[Иварне посуде и нерве
Снабдевање крви јајницима је због аа. Ет вв. Оварицае и утеринае. (. АА оварицае Дектра и синистра) оба јајника артерије протежу од предње површине аорте одмах испод бубрежних артерија, право најчешће потиче из аорте и лево - од бубрега артерије. Иде доле и бочно у односу на предњој површини слабински мишић, јајника артерија укршта сваки предњи уретер (која му гранчице), спољни илиац судове, као и граничну линију дође у карличне шупљине, које се налази овде у суспензорног лигамент јајника. Након медијално јајника артерија пролази између листова широке лигамента материце под јајоводима, дајући му гране, а затим - у мезентеријуму јајника; улази у капију јајника.
Гране оваријске артерије су широко анастомизоване са гранама јајника у матерничкој артерији. Веносни одлив из јајника врши се првенствено у венарском плексусу јајника, који се налази у пределу врата јајника. Стога одлив крви пролази у два правца: кроз утерине и јајнике. Права јајна вена има вентиле и улази у доњу вену. Лева јајоводна вена тече у леву бубрежну вену, без присуства вентила.
Лимфни одлив из јајника се јавља кроз лимфне посуде, нарочито обилне у пределу капија органа, где се излучује сублингуални лимфни плекус. Затим, лимф се преусмерава у пара-аортне лимфне чворове дуж лимфатских судова јајника.
Инхеренција јајника
Симпатичност - обезбеђује постганглионска влакна из целиаког (соларне), горње плетенице и хипогастричних плексуса; парасимпатички - због унутрашњих сакралних живаца.
Структура јајника
Површина је прекривена једним слојем јајника ембрионалних епитела. Испод тога налази се густо омотач везивног ткива (туница албугинеа). Везивно ткиво јајника образаца строме (строма оварии), богатих еластичних влакана. Садржај јајника, његовог паренхима, подељен је на спољашњи и унутрашњи слој. Унутрашњи слој, која се налази у центру јајника, ближе капије, назван срж (медулла оварии). У овом слоју, у лабаве везивног ткива су бројни крв и лимфни судови и нерви. Спољни слој јајника - оваријуми кортекса су густи. Има много везивног ткива у којима се налази сазревања примарне фоликула јајника (фоллицули оварици примарии), секундарна (Буббле) фоликула (фоллицули оварици сецундарии, с.весицулоси), а добро зреле фоликули граафови мехурићи (фоллицули оварици матурис), и жути и атретских тела.
У сваком фоликлу је женски репродуктивни јајум, или ооцит (овоцитус). Пречник ооците до 150 микрона, заобљених, садржи језгро, велику количину цитоплазме, који, поред мобилних органела, постоје протеин-липид инцлусионс (иолк), гликоген потребно за снабдевање јаја. Снабдевање ооцита обично потроши у року од 12-24 сата након овулације. Ако оплодња не дође, јаје умре.
Људски јајник има две мембране за покривање. Унутра је цитолемија, која је цитоплаземска мембрана ооцита. Иза цитолеме постоји слој такозваних фоликуларних ћелија које штите јаје и изводе функцију формирања хормона - ослобађају естрогене.
Физиолошки положај материце, тубуса и јајника обезбеђује апарат за суспендовање, причвршћивање и помоћ, Комбинујући перитонеум, лигаменте и целулозу карлице. Уређај за суспензију представљају упарене формације, обухвата округле и широке лигаменте материце, сопствене лигаменте и вијчане лигаментне јајнике. Широки лигамент материце, сопствени и суспендујући лигамент јајника чувају материцу у средњој позицији. Округли лигаменти антериорно привлаче дно материце и пружају физиолошки нагиб.
Уређај за причвршћивање осигурава положај шупљивог у средини мале карлице и чини га практично немогућим прелазити на бочне стране, напред и назад. Али пошто се лигаментни апарат одступа од материце у доњем делу, могуће је нагнути материцу у различитим правцима. До фиксирање уређаја подразумева и каблове распоређени у лабаве карлице ткиву и простиру од нижег матерични картице у бочном, предњих и задњих зидова карлице: сацроилиац магоцхние кардиналне, утерине и цистична-Весицо-стидна лигамента.
Поред месоварија, разликују се следећи лигаменти јајника:
- суспензија лигамента јајника, раније поменута као воронкотазоваиа. То је набор перитонеума са проширењем у њему крв (а. Ет в. Оварица) и лимфни судови и нерви јајника протезао између латералног карличног зида, лумбални фасције (ин дели заједничке бедрене артерије на спољашњег и унутрашњег) и горњи (цеви) крај јајника;
- лигамент личности оваријара пролази између листова широког матерњег лигамента, ближе задњем делу и спаја доњи крај јајника са бочном маргином материце. У материцу, лигамент овог јајника је причвршћен између почетка материце и кружног лигамента, напред и назад из другог. У дебљини лигамента су рр. Оварии, који су гранични прикључци матерничке артерије;
- апендикуларно-јајарски лигамент Клад се протеже око гребена апендикса на десни јајник или широк лигамент материце у облику гребена перитонеума. Лигамент је нестабилан и примећен је код 1/2 - 1/3 жена.
Пратећи апарат представљају мишићи и фасциа карличног пода, подељени на доњи, средњи и горњи (унутрашњи) слојеви.
Најснажнији је горњи (унутрашњи) слој мишића, који представља упарени мишић који подиже анус. Састоји се од мишићних снопова, који навијач из тртице до карлице у три правца (стидне-цоццигеал, Илио-цоццигеал и исцхиоцоццигеал мишића). Овај слој мишића се назива и дијафрагмом карлице.
Средњи слој мишића налази се између симфиза, стубних и исхијумских костију. Средњи слој мишића - урогенитална дијафрагма - заузима предњу половину карличног излаза, кроз њега пролази сечница и вагина. Између својим предњим летака су мишићни снопови који чине спољни сфинктер уретре, у задњи део постављених мишића снопове се протежу у попречном правцу, - тамно попречна Перинеална мишића.
Доњи (спољашњи) слој мишића дна карлице се састоји од површине која локација се у облику фигуре 8. Ово укључује булбоспонгиосус-цаверноус, исцхио-цаверноус, спољни сфинктер на ануса, површински попречни Перинеална мишића.
Онтогенеза јајника
Процес раста и атрезијом фоликула почиње са 20 недеља трудноће и на испоруку у девојку јајника је до 2 милиона јајне ћелије. У време менархеа, њихов број се смањује на 300 хиљада. Током читавог периода репродуктивног живота достиже зрелост и овулира не више од 500 фоликула. Почетни раст фоликула не зависи од стимулације ФСХ-а, ограничен је, а атресија се јавља брзо. Сматра се да су, уместо стероидних хормона, локални аутокринални / паракрине пептиди главни регулатор раста и атресија примарних фоликула. Сматра се да процес раста и атресија фоликула не прекидају никакви физиолошки процеси. Овај процес се наставља у свим годинама, укључујући интраутерини период и менопаузу, прекидан трудноћом, овулацијом и ановулацијом. Механизам који изазива раст фоликула и њихов број у сваком одређеном циклусу још увек није јасан.
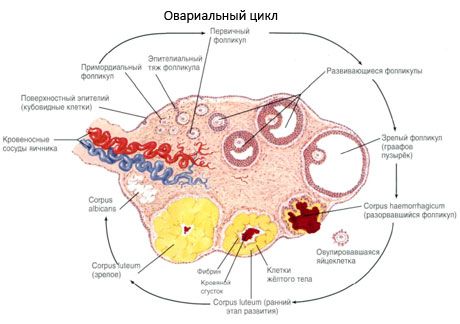
У свом развоју фоликул пролази кроз неколико фаза развоја. Примордијалне ћелијске ћелије пореклом из ендодерма жуманца, алантозе и мигрирају до гениталног подручја ембриона на 5. До 6. Недељу трудноће. Као резултат брзог митотског одељења, која траје од 6-8 недеља до 16-20 недеља трудноће, до 6 до 7 милиона ооцита се формира у јајницима ембриона, окружених танким слојем гранулозних ћелија.
Преантрални фоликул - ооцит је окружен мембраном (Зона пеллуцида). Гранулозне ћелије које окружују ооцит почињу да се пролиферишу, њихов раст зависи од гонадотропина и корелира са нивоом естрогена. Гранулоза ћелије су мета за ФСХ. У кораку преантрал фоликули гранулозно ћелије способне за синтезу три класе стероида: погодно изазива ароматазе активност, главни ензим који претвара андрогене у естрадиол. Верује се да естрадиол може повећати број својих рецептора, пружајући директан митогени ефекат на гранулозне ћелије независно од ФСХ. Сматра се као паракрине фактор који побољшава ефекте ФСХ, укључујући и активацију ароматизационих процеса.
ФСХ рецептори се појављују на мембранама гранулозних ћелија чим се појави раст фоликула. Редукција или повећање ФСХ доводи до промене у броју његових рецептора. Ова акција ФСХ модулира фактори раста. ФСХ делује кроз аденилатне-циклазе системом Г-протеина у фоликуле стероидогенезе иако углавном регулисано ФСХ, процес укључује многе факторе: јонске канале, рецептор тирозин киназе фосфолипазу систем секундарних гласника.
Улога андрогена у раном развоју фоликула је сложена. Гранулозне ћелије имају рецепторе за андрогене. Они нису подлогу само за ФСХ-индуковане ароматизације у естроген, али ниским концентрацијама могу побољшати процес арома. Када ниво андрогена повећава преантрал гранулозоцелуларни преференцијално ћелије означеном путу нема ароматизације у естрогене и једноставнији начин на замјену кроз андрогени 5а-редуктазе у андрогена развоју, који се не могу конвертовати у естроген, а тиме инхибирани ароматазе активност. Овај процес такође инхибира формирање ФСХ и ЛХ рецепторе, чиме зауставити развој фоликула.
Процес ароматизације, фоликула са високим нивоом андрогена пролази кроз процесе атресије. Раст и развој фоликла зависи од његове способности да претворе андрогене у естрогене.
У присуству ФСХ, доминантна супстанца фоликуларне течности је естрогена. У одсуству ФСХ - андрогена. ЛХ је нормална у фоликуларној течности до средине циклуса. Чим је повећан ниво митотску активности ЛХ гранулоза ћелија смањује, и дегенеративних промена јављају повишени ниво андрогена у нивоима фоликул стероида у фоллицулар течности него у плазми и одражава функционалну активност јајника: гранулозоцелуларни и тека ћелија. Ако су једини циљ за ФСХ гранулозне ћелије, онда ЛХ има много мете - то су ћелије, стромалне и лутеалне ћелије и гранулозне ћелије. Способност за стероидогенезу има и гранулозу и тека ћелије, али ароматазна активност превладава у гранулозним ћелијама.
Као одговор на ЛХ, ћелије тека производе андрогене, који се затим путем ароматизације индуковане ФСХ трансформишу гранулозним ћелијама у естрогене.
Као фоликула тека ћелија почињу да изразе гене за ЛХ рецептора П450 сец и 3бета-хидроксистероид дехидрогеназе, инсулину сличан фактор раста (ИГФ-1) у синергији са ЛХ повећања експресије гена, али не стимулишу стероидогенезу.
Оваријска стероидогенеза је увек зависна од ЛХ. Како се фоликул расте, тренутне ћелије изражавају ензим П450ц17, који формира андрогени из холестерола. Гранулозне ћелије немају овај ензим и зависе од тренутних ћелија у производњи естрогена из андрогена. За разлику од стероидогенезе - фоликулогенеза зависи од ФСХ. Као фоликула и повећати ниво естрогена долази у акциони повратне механизам - инхибира производњу ФСХ, што заузврат доводи до смањења ароматазе активности фоликула и, коначно фоликула атрезијом кроз апоптозу (програмирану ћелијску смрт).
Механизам повратних информација естрогена и ФСХ спречава развој фоликула који су почели да расте, али не и доминантни фоликул. Доминантни фоликул садржи више ФСХ рецептора који подржавају пролиферацију гранулозних ћелија и ароматизацију андрогена у естрогенима. Осим тога, паракрин и аутокринални пут дјелују као важан координатор за развој антралног фоликула.
Део аутокрино / паракрине регулатор су пептиди (инхибина, активин, фолистатин), гранулозно ћелије које се синтетишу као одговор на ФСХ и стигну у фоликуларне течности. Инхибин смањује секрецију ФСХ; активин стимулише ослобађање ФСХ од хипофизе и побољшава деловање ФСХ у јајнику; Фоллистатин потискује активност ФСХ, вероватно због везивања активина. Након овулације и развоја жутог тела, инхибит је под контролом ЛХ.
На раст и диференцијацију ћелија јајника утичу инсулин-лике фактори раста (ИГЕ). ИГФ-1 делује на гранулозне ћелије, што узрокује повећање цикличног аденозин монофосфата (цАМП), прогестерона, окситоцина, протеогликана и инхибита.
ИГФ-1 делује на тека ћелије, што доводи до повећања производње андрогена. Тека ћелије, заузврат, производе фактор туморске некрозе (ТНФ) и епидермални фактор раста (ЕГФ), који такође регулише ФСХ.
ЕГФ стимулише ширење гранулозних ћелија. ИГФ-2 је главни фактор раста фоликуларне течности, открио је и ИГФ-1, ТНФ-а, ТНФ-3 и ЕГФ.
Повреда паракрине и / или аутокрине регулације функције јајника изгледа да игра улогу у поремећајима овулационих процеса и формирању полицистичних јајника.
Како расте антрал фоликул, садржај естрогена у фоликуларној течности се повећава. На врхунцу њиховог пораста на гранулозним ћелијама појављују се рецептори за ЛХ, јавља се лутеинизација гранулозних ћелија, а производња прогестерона се повећава. Тако, током преовулационог периода, повећање производње естрогена узрокује појаву ЛХ рецептора, ЛХ, узрокује лутинизацију гранулозних ћелија и производњу прогестерона. Повећање прогестерона смањује ниво естрогена, који, очигледно, узрокује други врх ФСХ-а у средини циклуса.
Сматра се да овулација наступа 10-12 сати након врха ЛХ и 24-36 сати након врхунца естрадиола. Верује се да ЛХ стимулише смањење ооцита, лутеинизацију гранулозних ћелија, синтезу прогестерона и простагландина у фоликлу.
Прогестерон повећава активност протеолитичких ензима, заједно са простагландином који је укључен у рушење зида фоликула. ФСХ-индуковане прогестерона врхунац, омогућава излаз јајне ћелије из фоликула претварањем плазминогена на протеолитичку ензим - плазмин, обезбеђује довољну количину ЛХ рецептора за нормалан развој лутеалне фазе.
У року од 3 дана након овулације, гранулозним ћелијама повећава, појављују карактеристичне вакуолама испуњен пигмент - лутеин. Тека-лутеалне ћелије се разликују од теки и строма и постају део жутог тела. Врло брзо под утицајем ангиогенских фактора је развој капилара које прожимају цорпус лутеум, и побољшању васкуларизације повећао производњу естрогена и прогестерона. Активност стероидогенезе и животни век жутог тела одређује ниво ЛХ. Жуто тело није хомогени целуларни ентитет. Додавањем 2 врсте лутеална ћелија садржи ендотелне ћелије, макрофаге, фибробласти и друге. Велики лутеална ћелије продукују пептиде (Релакин, окситоцина) и да су активнији у Стероидогенесис још ароматазе активност и велике синтезе прогестерона од малих ћелија.
Највиши прогестерон се примећује осмог дана након врхунца ЛГ-а. Напоменуто је да се прогестерон и естрадиол у лутеалној фази спорадично излучују у корелацији са пулзним излазом ЛХ. Са формирањем жутог тела, контрола над производњом инхибитних пролаза од ФСХ до ЛХ. Инхибин је повећана са порастом естрадиола до врха ЛХ и настављају да расту након пика ЛХ, мада нивои естрогена смањује. Иако су инхибит и естрадиол излучени гранулозним ћелијама, регулишу се на различите начине. Смањење инхибина на крају лутеалне фазе доприноси повећању ФСХ за следећи циклус.
Жуто тело веома брзо - на 9-11 дана после смањења овулације.
Механизам дегенерације није јасан и није повезан са лутеолитичком улогом естрогена или са механизмом повезан са рецептором, као што се види у ендометрију. Постоји још једно објашњење за улогу естрогена произведених од жутог тела. Познато је да су за синтезу прогестеронских рецептора у ендометрију потребна естрогена. Естрогени лутеалне фазе су вероватно неопходни за прогестеронске промене у ендометрију после овулације. Неадекватан развој прогестеронских рецептора, као резултат неадекватног садржаја естрогена, вероватно је додатни механизам неплодности и губитка ране трудноће, још један облик инфериорности лутеалне фазе. Сматра се да је животни век жутог тела постављен у време овулације. И сигурно ће се регресирати ако хорионски гонадотропин није подржан у вези са трудноћом. Према томе, регресија жутог тела доводи до смањења нивоа естрадиола, прогестерона и инхибина. Редукцијски инхибитор уклања свој инхибиторни ефекат на ФСХ; смањење естрадиола и прогестерона омогућава врло брзо да обнови секрецију ГнРХ и уклони механизам повратне спреге од хипофизе. Редукција инхибина и естрадиола, заједно са повећањем ГнРХ, доводи до преваленције ФСХ у односу на ЛХ. Повећање ФСХ доводи до раста фоликула уз накнадни избор доминантног фоликула, а почиње нови циклус у случају да се трудноћа не појави. Стероидни хормони играју водећу улогу у репродуктивној биологији иуопште у физиологији. Одређују фенотип особе, утичу на кардиоваскуларни систем, метаболизам костију, кожу, опште добро тело и играју кључну улогу у трудноћи. Акција стероидних хормона одражава интрацелуларне и генетске механизме који су неопходни за преношење екстрацелуларног сигнала на језгро ћелије како би изазвали физиолошки одговор.
Естрогени дифузују кроз ћелијску мембрану и везују се за рецепторе који се налазе у језгру ћелије. Комплекс рецептора-стероида се затим везује за ДНК. У циљним ћелијама, ове интеракције воде ка експресији гена, синтези протеина, специфичној функцији ћелија и ткива.