
Синтеза, секрецење и метаболизам катехоламина
Последње прегледано: 19.10.2021

Сви иЛиве садржаји су медицински прегледани или проверени како би се осигурала што већа тачност.
Имамо стриктне смјернице за набавку и само линкамо на угледне медијске странице, академске истраживачке институције и, кад год је то могуће, медицински прегледане студије. Имајте на уму да су бројеви у заградама ([1], [2], итд.) Везе које се могу кликнути на ове студије.
Ако сматрате да је било који од наших садржаја нетачан, застарио или на неки други начин упитан, одаберите га и притисните Цтрл + Ентер.
Церебрални слој надбубрежних жлезда производи једињење далеко од стероида структуре. Они садрже језгро 3,4-дихидроксифенила (катехол) и називају се катехоламини. То укључује адреналин, норепинефрин и допамин бета-окси-тирин.
Синтеза Секуенце катехоламина је веома једноставан: Тирозин → дихидрокипхенилаланине (ДОПА) → → допамин норепинефрин → адреналин. Тирозин улази у организам храном, али може се формирати из фенилаланина у јетри под дејством фенилаланина хидроксилазе. Крајња реакциона производи тирозина у различитим ткивима. У адреналне мождине процес приступа се фази формирања адреналина, на крајевима симпатичких нерава - норадреналин, у одређеним неуронима централног нервног система катехоламина допамин синтезе попуњеног обрасца.
Претварање тирозина у ДОПА катализује тирозин хидроксилаза, чији су кофактори тетрахидро-биоптерин и кисеоник. Верује се да је овај ензим који ограничава брзину читавог процеса биосинтезе катехоламина и да је спречен крајњим производима процеса. Тирозин хидроксилаза је главни циљ регулаторних ефеката на биосинтезу катехоламина.
Допа то допамина конверзија је катализована ензимом ДОПА-декарбоксилазе (кофактора - пиридоксал) који је релативно неспецифичне и декарбоксилована и други ароматични Л-амино киселине. Међутим, постоје индикације могућности модификације синтезе катехоламина промјеном активности и овог ензима. У неким неуронима нема ензима за даљу конверзију допамина, и то је крајњи производ. Других ткива садрже допамин-бета-хидроксилазе (кофакторе - бакар, аскорбинске киселине и кисеоника) која конвертује допамин то норепинефрин. У надбубрежне сржи (али не симпатички нервни завршеци) присутна пхенилетханоламине - метхилтрансферасе формирање адреналин из норадреналина. Донатор метил групе у овом случају је С-аденозилметионин.
Важно је запамтити да синтеза пхенилетханоламине-Н-Метилтрансферази изазвана глукокортикоида који спадају у церебралном кортикалном слоју портала венског система. Ово може објаснити чињенице огледа комбинују два различита ендокриних жлезда у једном телу. Значење глукокортикоидног синтезе адреналина наглашен чињеницом да су адреналне мождине ћелије производећи норепинефрин, пореданих око артеријских судова, док крвне ћелије су добијене адреналинпродутсируиусцхие суштини венских синуса, локализован у надбубрега кортексу.
Колапс катехоламина настаје углавном под утицајем два ензима система: цатецхол-О-метилтрансфераза (ЦОМТ) и моноамин оксидазе (МАО). Главни начини епинефрина и норепинефрина распадања је шематски приказано на слици. 54. Под дејством ЦОМТ у присуству донатора метил група С-адренозилметионина катехоламина и претвара у норметанфрин метанефрин (3-О-метил деривата епинефрина и норепинефрина), који под утицајем МАО претворен алдехида и више (у присуству алдехида) у ваниллил-бадемова киселина (ИЦХ) - главни производ разлагања норадреналин и адреналина. У истом предмету, када први изложен катехоламина МАО акцију, не ЦОМТ, они су конвертовани у 3,4-диоксиминдалеви алдехида, а затим под утицајем алдехида и ЦОМТ - 3,4-диоксиминдалнуиу киселине и ИУД. У присуству алцохол дехидрогенасе катехоламина могу формирати 3-метокси-4-оксифенилгликол, главни крајњег производа деградације епинефрина и норепинефрина у ЦНС.
Дисинтегратион допамин наставља слично, осим да његове метаболити су лишени хидроксил група на атому бета-угљеника и стога, уместо ваниллил-бадемова киселина формирани хомованиллиц (ХВА) и 3-метокси-4-оксифенилуксуснаиа киселине.
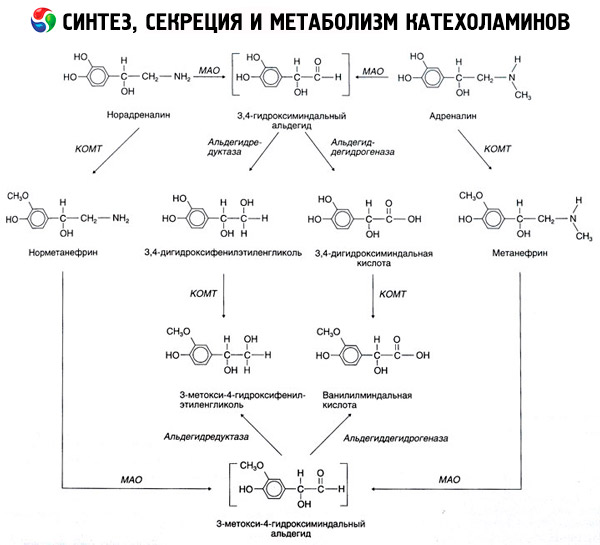
Постојање киноидног пута за оксидацију молекула катехоламина, на коме се могу постулирати и интермедијарни производи са израженом биолошком активношћу.
Формиран деловањем цитосолне ензима, адреналина и норадреналина на симпатичним нервним завршецима, адреналне мождине и унесите секреторне грануле које их штити од деловања деградативни ензим. Заузимање катехоламина са гранулама захтијева трошкове енергије. У хромафинском гранулама на адреналне мождине катехоламина чврсто везаних за АТП (у односу 4: 1) и специфични протеини - Цхромогранин који спречава дифузију хормона из гранула у цитоплазми.
Директна подстицај излучивања катехоламина очигледно пенетрације калцијум целл стимулисање егзоцитозу (Фусион мембране гранула ћелијској површини и њихову јаза са укупним приносом од растворљивих садржаја - катехоламина, допамин-бета-хидроксилазе, АТП и Цхромогранин - у екстрацелуларне течности) .
Физиолошки ефекти катехоламина и механизам њихове акције
Ефекти катехоламина почињу интеракцијом са специфичним рецепторима циљних ћелија. Ако се рецептори тироидних и стероидних хормона локализован унутар ћелија, Катехоламински рецептори (као ацетилхолина и пептидни хормони) су присутне на спољашњој површини ћелије.
Одавно је утврђено да у погледу неке реакције адреналина или норадреналина су ефикаснији од синтетичког катехоламина изопротеренола, док за друге ефекат супериоран деловању изопротеренола епинефрина или норепинефрин. На основу тога развијен је концепт присуства у ткивима две врсте адренорецептора: алфа и бета, ау неким од њих постоји само један од ова два типа. Изопротеренол је најмоћнија агонист бета-адренергичких рецептора, док синтетичке једињење фенилефрином - најмоћнији агонист алфа-адренергичких рецептора. Натурал катехоламина - адреналина и норадреналина - могу за интеракцију са рецепторима оба типа, али адреналин узима већи афинитет за бета, и норепинефрина - алфа-рецептори.
Катехоламина јаче активирање срчане бета-адренергички рецептори него бета рецепторе глатких мишића, омогућавајући бета-типе подељен у подтипове: бета1 рецепторе (срца, масне ћелије) и бета2 рецептора (бронха, крвних судова, итд ...). Деловање изопротеренола на бета1 рецептора супериорном дејством адреналина и норадреналина само 10 пута, док су бета2-рецепторе делује 100-1000 пута снажнији од природног катехоламина.
Специфични антагонисти дјубрива (феноксибензамин и фентоламин против алфа- и пропранолол против бета-рецептора) потврдила адекватност класификације адреноцепторима. Допамина може да интерагује са оба алфа- и бета рецептора, већ у различитим ткивима (мозак, хипофиза, судова) пронађено и сопствени допаминергијски рецепторе специфичне блокера који је халоперидол. Број бета рецептора се креће од 1000 до 2000 по ћелији. Биолошки ефекти катехоламина посредованих бета-рецептора су повезани, по правилу, са активацијом аденилат циклазе и повећане интрацелуларног цАМП. Рецептор и ензим, иако су функционално повезани, али представљају различите макромолекуле. Модулација аденилатне циклазе под утицајем комплекса хормона-рецептор су укључени гуанозин трифосфат (ГТП) и друге пурин нуклеотиди. Повећавајући активност ензима, чини се да смањују афинитет бета рецептора за агонисте.
Феномен повећања сензитивности денервираних структура одавно је познат. Насупрот томе, продужена изложеност агонистима смањује осетљивост циљних ткива. Истраживање бета рецептора омогућило је да објасне ове појаве. Показано је да продужено дејство изопротеренола доводи до губитка осетљивости аденилат циклазе због смањења броја бета рецептора.
Процес десензитизације не захтева активацију синтезе протеина и вероватно је последица постепеног стварања иреверзибилних комплекса хормонских рецептора. Напротив, примена 6-оксидофамина, која разбија симпатичке крајеве, прати повећање броја реакција бета рецептора у ткивима. Није искључено да повећање симпатичног нервног деловања одређује десенситизацију крвних судова и масног ткива у односу на узрасту у односу на катехоламине.
Број адренорецептора у различитим органима може контролисати други хормони. Стога, повећава естрадиол, прогестерон и смањити број алфа-адренергичких рецептора у материци, који је праћен одговарајућим повећању и смањењу њеног контрактилног одговора на катехоламина. Ако интрацелуларног "други месенгер", формиран деловањем агониста бета-рецептора, свакако је цАМП, у односу на предајник алфа-адренергичких ефеката компликованија. Претпоставља се да постоје различити механизми: смањење нивоа цАМП-а, повећање садржаја цАМП-а, модулација ћелијске динамике калцијума,
За репродукцију различитих ефеката у телу, обично су потребне дозе епинефрина, које су 5-10 пута ниже од норепинефрина. Мада се ефикаснија у односу на а- и бета1-адренергични рецептори, важно је запамтити да и ендогена катехоламина може да комуницира са оба алфа- и бета рецептора. Стога, биолошки одговор овог тела на адренергичну активацију у великој мери зависи од врсте рецептора присутних у њему. Међутим, то не значи да је селективно активирање нервне или хуморалне везе симпатичко-надбубрежног система немогуће. У већини случајева постоји интензивирана активност различитих веза. Стога се претпоставља да активира рефлекс хипогликемије адреналне мождине, док је смањење крвног притиска (држању хипотензија) праћена углавном норепинефрина ослобађање из симпатичним нервним завршецима.
Адренорецептори и ефекти њихове активације у различитим ткивима
|
Систем, орган |
Адреноцептор тип |
Реакција |
|
Кардиоваскуларни систем: |
||
|
Срце |
Бета |
Повећање фреквенције контракција, проводљивости и контрактилности |
|
Артериоли: |
||
|
Кожу и мукозне мембране |
Алфа |
Смањење |
|
Скелетних мишића |
Бета |
Редукција продужења |
|
Абдоминални органи |
Алфа (више) |
Смањење |
|
Бета |
Проширење |
|
|
Вене |
Алфа |
Смањење |
|
Респираторни систем: |
||
|
Бронхијални мишићи |
Бета |
Проширење |
|
Систем за варење: |
||
|
Стомак |
Бета |
Смањена функција мотора |
|
Црева |
Алфа |
Смањење сфинктера |
|
Сплеен |
Алфа |
Смањење |
|
Бета |
Релаксација |
|
|
Вањски тајни део панкреаса |
Алфа |
Смањено лучење |
|
Генитоуринарни систем: |
Алфа |
Снижење сфинктера |
|
Бешика |
Бета |
Опуштање егзорцистичког мишића |
|
Мушки сексуални органи |
Алфа |
Ејакулација |
|
Очи |
Алфа |
Ученик је дилиран |
|
Кожа |
Алфа |
Повећано знојење |
|
Пљувачке жлезде |
Алфа |
Изолација калијума и воде |
|
Бета |
Секретирање амилазе |
|
|
Ендокрине жлезде: |
||
|
Отоци панкреаса |
||
|
Бета ћелије |
Алфа (више) |
Смањено инсулинско лучење |
|
Бета |
Повећана секреција инсулина |
|
|
Алфа ћелије |
Бета |
Повећана секреција глукагона |
|
8-ћелија |
Бета |
Повећана секреција соматостатина |
|
Хипоталамус и хипофиза: |
||
|
Соматотрофи |
Алфа |
Повећана секреција СТХ |
|
Бета |
Смањена секреција СТХ |
|
|
Лактотрофи |
Алфа |
Смањено лучење пролактина |
|
Тиротрофи |
Алфа |
Смањено лучење ТСХ |
|
Кортикотрофи |
Алфа |
Повећана секреција АЦТХ |
| бета | Смањена секреција АЦТХ | |
|
Штитна жлезда: |
||
|
Фоликуларне ћелије |
Алфа |
Смањено лучење тироксина |
|
Бета |
Повећана секреција тироксина |
|
|
Парафоликуларне (К) ћелије |
Бета |
Повећана секреција калцитонина |
|
Паратироидне жлезде |
Бета |
Повећана секреција ПТХ |
|
Бубрези |
Бета |
Повећана секреција ренина |
|
Стомак |
Бета |
Повећати секрецију гастрина |
|
Основна размена |
Бета |
Повећање потрошње кисеоника |
|
Јетра |
? |
Повећање гликогенолизе и глуконеогенезе из приноса глукозе; повећати кетогенезу са ослобађањем кетонских тијела |
|
Адипозно ткиво |
Бета |
Повећање липолизе са ослобађањем слободних масних киселина и глицерола |
|
Скелетни мишићи |
Бета |
Повећање гликолизе са ослобађањем пирувата и лактата; смањење протеолизе са смањењем приноса аланина, глутамина |
Важно је имати на уму да су резултати интравенске примене катехоламина нису увек адекватно одражавају ефекте ендогених једињења. Ово се односи углавном на норадреналина, јер стоји у телу углавном није крв, али директно у синаптичке пукотине. Стога ендогени норепинефрин активира, на пример, није само васкуларни алфа рецептори (повишен крвни притисак), већ и срца бета-рецепторе (лупање срца), док администрацију норадреналина спољних доводи углавном до активације васкуларног алфа рецептора и рефлекса (преко вагус) успоравање палпитације.
Ниске дозе епинефрина активирају углавном бета рецепторе мишићних судова и срца, што доводи до пада периферног васкуларног отпора и повећања минутног волумена срца. У неким случајевима, први ефекат може доминирати, а након примене адреналина, развија се хипотензија. У већим дозама адреналин такође активира алфа рецепторе, што је праћено повећањем периферне васкуларне резистенције и на позадини повећања тренутног волумена срца доводи до повећања крвног притиска. Међутим, његов учинак на васкуларним бета рецепторима је такође сачуван. Као резултат, повећање систолног притиска превазилази сличну вредност дијастолног притиска (повећање импулзног притиска). Уз увођење још већих доза, почињу превладати алфа-миметички ефекти епинефрина: систолни и дијастолни притисак се повећава паралелно, и под утицајем норадреналина.
Ефекат катехоламина на метаболизам се састоји од њихових директних и индиректних ефеката. Први се реализују углавном путем бета-рецептора. Сложени процеси су повезани са јетром. Иако се повећање хепатичне гликогенолизе традиционално сматра резултатом активације бета рецептора, постоје и подаци о укључивању алфа рецептора у ово. Посредовани ефекти катехоламина су повезани са модулацијом секреције многих других хормона, на пример инсулина. У дјеловању адреналина на његову секрецију, алфа-адренергична компонента јасно доминира, пошто се показује да било који стрес прати инхибицијом секреције инсулина.
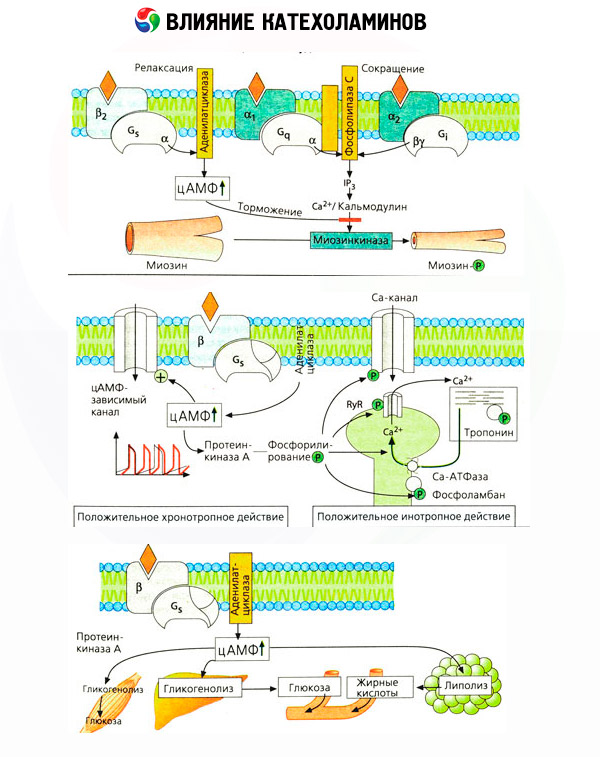
Комбинација директних и индиректних ефеката катехоламина доводи до хипергликемије, коњугат не само са повећањем производњу глукозе у јетри, али и са инхибицијом његовог коришћења од стране периферних ткива. Убрзање липолизе узрокује хиперлипацидемију са повећаном испоруком масних киселина у јетру и интензивирање производње кетонских тијела. Јачање гликолизе у мишићима доводи до повећања производње у млечне киселине у крви и пирувата, која заједно са глицерола ослобођеног из масног ткива, су претходници у глукогенезом јетре.
Регулација секреције катехоламина. Сличност производа и метода реаговања симпатичког нервног система и адреналне мождине била је основа за комбиновање ових структура у један тело симпатхоадренал систем ослобађања од живчаног и хормоналног питању веза. Различити аферентна сигнали се концентришу у хипоталамусу и центара кичмене мождине и продужену мождину од којих потичу ефферент парцела Укључивање преганглиониц неурон целл тела налазе у латералном рога кичмене мождине на нивоу цервикалне ВИИИ - ИИ-ИИИ лумбалном сегментима.
Преганглиониц аксона ових ћелија напуштају кичмену мождину и формира синаптичких веза са неурони су локализовани у ганглији симпатичког ланца, или адреналне мождине ћелија. Ово су холинергични преганглиониц влакна. Први фундаментални Разлика постганглијским симпатетичких неурона и надбубрежне сржи ћелије хромафина састоји у да други се преноси долазећег сигнала је холинергични неуро проводљивост (постганглијским адренергични живце) и хуморални истицањем адренергички једињења у крви. Друга разлика се своди на постганглијским нерава који производе норепинефрин, док су адреналне мождине ћелија - пожељно адреналина. Ове две супстанце имају различите ефекте на тканини.