Медицински стручњак за чланак

Нове публикације
хипофиза
Последње прегледано: 04.07.2025

Сви иЛиве садржаји су медицински прегледани или проверени како би се осигурала што већа тачност.
Имамо стриктне смјернице за набавку и само линкамо на угледне медијске странице, академске истраживачке институције и, кад год је то могуће, медицински прегледане студије. Имајте на уму да су бројеви у заградама ([1], [2], итд.) Везе које се могу кликнути на ове студије.
Ако сматрате да је било који од наших садржаја нетачан, застарио или на неки други начин упитан, одаберите га и притисните Цтрл + Ентер.
Хипофиза (hypophysis, s.glandula pituitaria) се налази у хипофизној јами sella turcica сфеноидне кости и одвојена је од лобање наставком dura mater мозга, формирајући дијафрагму sella. Кроз отвор у овој дијафрагми, хипофиза је повезана са инфундибулумом хипоталамуса диенцефалона. Попречна величина хипофизе је 10-17 mm, антеропостериорна 5-15 mm, вертикална 5-10 mm. Маса хипофизе код мушкараца је приближно 0,5 g, код жена - 0,6 g. Хипофиза је споља покривена капсулом.
У складу са развојем хипофизе из два различита зачатка, у органу се разликују два режња - предњи и задњи. Аденохипофиза, или предњи режањ (adenohypophysis, s.lobus anterior), је већи и чини 70-80% укупне масе хипофизе. Гушћи је од задњег режња. У предњем режњу се разликују дистални део (pars distalis), који заузима предњи део хипофизне јаме, средњи део (pars intermedia), који се налази на граници са задњим режњем, и гомољасти део (pars tuberalis), који иде навише и повезује се са инфундибулумом хипоталамуса. Због обиља крвних судова, предњи режањ има бледожуту боју са црвенкастим нијансом. Паренхим предњег дела хипофизе представљен је са неколико врста жлезданих ћелија, између чијих се нити налазе синусоидни крвни капилари. Половина (50%) ћелија аденохипофизе су хромофилни аденоцити, који у својој цитоплазми имају ситнозрнасте грануле које се добро боје солима хрома. То су ацидофилни аденоцити (40% свих ћелија аденохипофизе) и базофилни аденоцити (10%). Базофилни аденоцити укључују гонадотропне, кортикотропне и тиротропне ендокриноците. Хромофобни аденоцити су мали, имају велико једро и малу количину цитоплазме. Ове ћелије се сматрају прекурсорима хромофилних аденоцита. Осталих 50% ћелија аденохипофизе су хромофобни аденоцити.
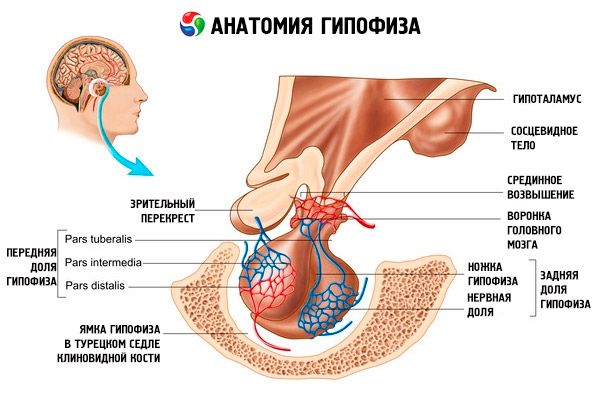
Неурохипофиза, или задњи режањ (neurohypophysis, s.lobus posterior), састоји се од нервног режња (lobus nervosus), који се налази у задњем делу хипофизне јаме, и левка (infundibulum), који се налази иза туберозног дела аденохипофизе. Задњи режањ хипофизе формирају неуроглијалне ћелије (ћелије хипофизе), нервна влакна која иду од неуросекреторних језгара хипоталамуса до неурохипофизе и неуросекреторна телашца.
Хипофиза је, преко нервних влакана (путева) и крвних судова, функционално повезана са хипоталамусом диенцефалона, који регулише активност хипофизе. Хипофиза и хипоталамус, заједно са својим неуроендокриним, васкуларним и нервним везама, обично се сматрају хипоталамус-хипофизним системом.
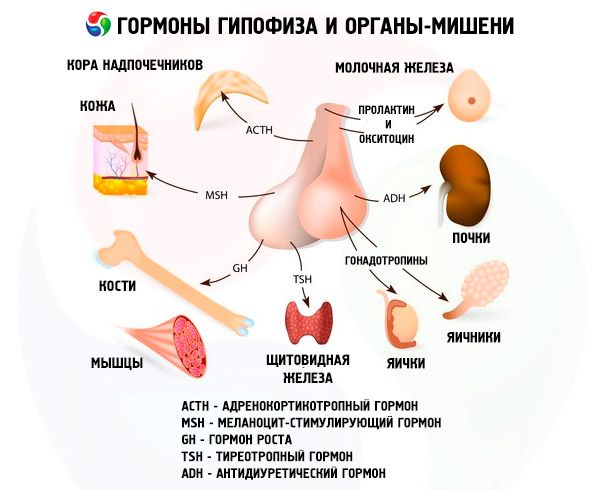
Хормони предње и задње хипофизе утичу на многе функције тела, првенствено преко других ендокриних жлезда. У предњој хипофизи, ацидофилни аденоцити (алфа ћелије) производе соматотропни хормон (хормон раста), који учествује у регулацији процеса раста и развоја код младих организама. Кортикотропни ендокриноцити луче адренокортикотропни хормон (АЦТХ), који стимулише лучење стероидних хормона надбубрежних жлезда. Тиротропни ендокриноцити луче тиротропни хормон (ТСХ), који утиче на развој штитне жлезде и активира производњу њених хормона. Гонадотропни хормони: фоликулостимулишући (ФСХ), лутеинизирајући (ЛХ) и пролактин - утичу на полно сазревање тела, регулишу и стимулишу развој фоликула у јајнику, овулацију, раст млечних жлезда и производњу млека код жена, процес сперматогенезе код мушкараца. Ове хормоне производе базофилни аденоцити (бета ћелије). Овде се луче и липотропни фактори хипофизе, који утичу на мобилизацију и коришћење масти у организму. У средњем делу предњег режња формира се меланоцит-стимулирајући хормон, који контролише стварање пигмената - меланина - у телу.
Неуросекреторне ћелије супраоптичког и паравентрикуларног једра у хипоталамусу производе вазопресин и окситоцин. Ови хормони се транспортују до ћелија задњег режња хипофизе дуж аксона који чине хипоталамо-хипофизни тракт. Из задњег режња хипофизе ове супстанце улазе у крв. Хормон вазопресин има вазоконстриктивно и антидиуретско дејство, због чега се назива и антидиуретички хормон (АДХ). Окситоцин има стимулативно дејство на контрактилност мишића материце, повећава лучење млека млечном жлездом која лактира, инхибира развој и функцију жутог тела и утиче на промене тонуса глатких (непругастих) мишића гастроинтестиналног тракта.
Развој хипофизе
Предњи режањ хипофизе развија се из епитела дорзалног зида усног залива у облику прстенастог израстања (Раткеова кесица). Ово ектодермално избочење расте према дну будуће треће коморе. Према њему, са доње површине друге мождане везикуле (будуће дно треће коморе), расте наставак, из којег се развијају сиви туберкулум инфундибулума и задњи режањ хипофизе.
Крвни судови и живци хипофизе
Из унутрашњих каротидних артерија и крвних судова артеријског круга великог мозга, горња и доња хипофизна артерија усмеравају се ка хипофизи. Горње хипофизне артерије иду ка сивом једру и инфундибулуму хипоталамуса, овде анастомозирају једна са другом и формирају капиларе који продиру у мождано ткиво - примарну хемокапиларну мрежу. Из дугих и кратких петљи ове мреже формирају се порталне вене, које су усмерене ка предњем режњу хипофизе. У паренхиму предњег режња хипофизе, ове вене се разлажу у широке синусоидне капиларе, формирајући секундарну хемокапиларну мрежу. Задњи режањ хипофизе снабдева се крвљу углавном доњом хипофизном артеријом. Између горње и доње хипофизне артерије постоје дугачке артеријске анастомозе. Одлив венске крви из секундарне хемокапиларне мреже врши се кроз систем вена које се уливају у кавернозне и интеркавернозне синусе дуре матер мозга.
Симпатичка влакна која улазе у орган заједно са артеријама учествују у инервацији хипофизе. Постганглијска симпатичка нервна влакна одлазе из плексуса унутрашње каротидне артерије. Поред тога, у задњем режњу хипофизе налазе се бројни завршеци процеса неуросекреторних ћелија које се налазе у језгрима хипоталамуса.
Карактеристике хипофизе повезане са старењем
Просечна тежина хипофизе код новорођенчади достиже 0,12 г. Тежина органа се удвостручује до 10. године, а утростручује до 15. године. До 20. године тежина хипофизе достиже свој максимум (530-560 мг) и остаје готово непромењена у наредним узрасним периодима. Након 60 година, примећује се благо смањење тежине ове ендокрине жлезде.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Хормони хипофизе
Јединство нервне и хормонске регулације у телу обезбеђено је блиском анатомском и функционалном везом хипофизе и хипоталамуса. Овај комплекс одређује стање и функционисање целог ендокриног система.
Главна ендокрина жлезда која производи низ пептидних хормона који директно регулишу функцију периферних жлезда је хипофиза. То је црвенкасто-сива формација у облику пасуља прекривена фиброзном капсулом тежине 0,5-0,6 г. Она се незнатно разликује у зависности од пола и старости особе. Општеприхваћена подела хипофизе на два режња која се разликују по развоју, структури и функцији остаје: предњи дистални - аденохипофиза и задњи - неурохипофиза. Први чини око 70% укупне масе жлезде и конвенционално се дели на дистални, инфундибуларни и средњи део, други - на задњи део, или режањ, и стабљику хипофизе. Жлезда се налази у хипофизној јами турског седла сфеноидне кости и повезана је са мозгом преко стабљике. Горњи део предњег режња прекривен је оптичком хијазмом и оптичким трактовима. Снабдевање крвљу хипофизе је веома обилно и врши се гранама унутрашње каротидне артерије (горња и доња хипофизна артерија), као и гранама артеријског круга великог мозга. Горње хипофизне артерије учествују у снабдевању крвљу аденохипофизе, а доње - неурохипофизе, додирујући се са неуросекреторним завршецима аксона великих ћелијских једара хипоталамуса. Прве улазе у средњу еминенцију хипоталамуса, где се распршују у капиларну мрежу (примарни капиларни плексус). Ови капилари (са којима се контактирају завршеци аксона малих неуросекреторних ћелија медиобазалног хипоталамуса) сакупљају се у порталне вене које се спуштају дуж стабљике хипофизе у паренхим аденохипофизе, где се поново деле у мрежу синусоидних капилара (секундарни капиларни плексус). Дакле, крв, након што је претходно прошла кроз средњу еминенцију хипоталамуса, где је обогаћена хипоталамичким аденохипофизотропним хормонима (ослобађајућим хормонима), улази у аденохипофизу.
Одлив крви засићене аденохипофизним хормонима из бројних капилара секундарног плексуса врши се кроз систем вена, које се затим уливају у венске синусе дуре матер, а затим у општи крвоток. Дакле, портални систем хипофизе са силазним смером протока крви из хипоталамуса је морфофункционална компонента сложеног механизма неурохуморалне контроле тропских функција аденохипофизе.
Хипофизу инервишу симпатичка влакна која прате хипофизне артерије. Она потичу од постганглијских влакана која пролазе кроз унутрашњи каротидни плексус, повезан са горњим цервикалним ганглијама. Не постоји директна инервација аденохипофизе из хипоталамуса. Задњи режањ прима нервна влакна из неуросекреторних језгара хипоталамуса.
Аденохипофиза је веома сложена формација у својој хистолошкој архитектури. Има две врсте жлезданих ћелија - хромофобне и хромофилне. Потоње се пак деле на ацидофилне и базофилне (детаљан хистолошки опис хипофизе дат је у одговарајућем одељку приручника). Међутим, треба напоменути да су хормони које производе жлездане ћелије које чине паренхим аденохипофизе, због разноликости потоњих, донекле различити по својој хемијској природи, а фина структура ћелија које секретују мора одговарати карактеристикама биосинтезе сваке од њих. Али понекад се у аденохипофизи могу приметити прелазни облици жлезданих ћелија које су способне да производе неколико хормона. Постоје докази да тип жлезданих ћелија аденохипофизе није увек генетски одређен.
Испод дијафрагме турског седла налази се левкасти део предњег режња. Он обухвата дршку хипофизе, додирујући сиви туберкулум. Овај део аденохипофизе карактерише присуство епителних ћелија и обилна снабдевање крвљу. Такође је хормонски активан.
Средњи (интермедијарни) део хипофизе састоји се од неколико слојева великих секреторно-активних базофилних ћелија.
Хипофиза обавља разне функције путем својих хормона. Њен предњи режањ производи адренокортикотропни (АКТХ), тиреостимулирајуће (ТСХ), фоликулостимулирајуће (ФСХ), лутеинизирајуће (ЛХ), липотропне хормоне, као и хормон раста - соматотропни (СТО) и пролактин. У средњем режњу се синтетише меланоцитстимулирајући хормон (МСХ), а у задњем режњу се акумулирају вазопресин и окситоцин.
АЦТХ
Хормони хипофизе су група протеинских и пептидних хормона и гликопротеина. Од хормона предњег режња хипофизе, АЦТХ је најбоље проучен. Производе га базофилне ћелије. Његова главна физиолошка функција је стимулација биосинтезе и секреције стероидних хормона од стране коре надбубрежне жлезде. АЦТХ такође показује меланоцит-стимулишућу и липотропну активност. Године 1953. изолован је у чистом облику. Касније је утврђена његова хемијска структура, која се састоји од 39 аминокиселинских остатака код људи и бројних сисара. АЦТХ нема специфичност за врсту. Тренутно је спроведена хемијска синтеза и самог хормона и различитих фрагмената његовог молекула, активнијих од природних хормона. Структура хормона има два дела пептидног ланца, од којих један обезбеђује детекцију и везивање АЦТХ за рецептор, а други пружа биолошки ефекат. Очигледно се везује за АЦТХ рецептор кроз интеракцију електричних наелектрисања хормона и рецептора. Улогу биолошког ефектора АЦТХ обавља фрагмент молекула 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
Меланоцит-стимулишућа активност АЦТХ је последица присуства у молекулу Н-терминалног региона, који се састоји од 13 аминокиселинских остатака и понавља структуру алфа-меланоцит-стимулишућег хормона. Овај регион такође садржи хептапептид присутан у другим хипофизним хормонима и поседује нека адренокортикотропна, меланоцит-стимулишућа и липотропна дејства.
Кључним тренутком у деловању АЦТХ сматра се активација ензима протеин киназе у цитоплазми уз учешће цАМП-а. Фосфорилована протеин киназа активира ензим естеразу, која претвара естре холестерола у слободну супстанцу у капљицама масти. Протеин синтетисан у цитоплазми као резултат фосфорилације рибозома стимулише везивање слободног холестерола за цитохром П-450 и његов пренос из капљица липида у митохондрије, где су присутни сви ензими који обезбеђују конверзију холестерола у кортикостероиде.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Хормон који стимулише штитну жлезду
ТСХ - тиротропин - главни регулатор развоја и функционисања штитне жлезде, процеса синтезе и секреције тироидних хормона. Овај сложени протеин - гликопротеин - састоји се од алфа и бета подјединица. Структура прве подјединице се поклапа са алфа подјединицом лутеинизујућег хормона. Штавише, у великој мери се поклапа код различитих животињских врста. Секвенца аминокиселинских остатака у бета подјединици људског ТСХ је дешифрована и састоји се од 119 аминокиселинских остатака. Може се приметити да су бета подјединице људског и говеђег ТСХ сличне у много чему. Биолошка својства и природа биолошке активности гликопротеинских хормона одређени су бета подјединицом. Она такође обезбеђује интеракцију хормона са рецепторима у различитим циљним органима. Међутим, бета подјединица код већине животиња показује специфичну активност тек након што се комбинује са алфа подјединицом, која делује као нека врста активатора хормона. Потоња са једнаком вероватноћом индукује лутеинизујуће, фоликулостимулишуће и тиротропне активности одређене својствима бета подјединице. Пронађена сличност нам омогућава да закључимо да су ови хормони настали у процесу еволуције од једног заједничког претходника, бета подјединица такође одређује имунолошка својства хормона. Постоји претпоставка да алфа подјединица штити бета подјединицу од дејства протеолитичких ензима, а такође олакшава њен транспорт из хипофизе до периферних циљних органа.
Гонадотропни хормони
Гонадотропини су присутни у телу у облику ЛХ и ФСХ. Функционална сврха ових хормона се генерално своди на обезбеђивање репродуктивних процеса код јединки оба пола. Они су, као и ТСХ, сложени протеини - гликопротеини. ФСХ индукује сазревање фоликула у јајницима код жена и стимулише сперматогенезу код мушкараца. ЛХ изазива пуцање фоликула код жена са формирањем жутог тела и стимулише лучење естрогена и прогестерона. Код мушкараца исти хормон убрзава развој интерстицијалног ткива и лучење андрогена. Ефекти гонадотропина су међусобно зависни и јављају се синхроно.
Динамика секреције гонадотропина код жена се мења током менструалног циклуса и довољно је детаљно проучена. У преовулаторној (фоликуларној) фази циклуса, садржај ЛХ је на прилично ниском нивоу, док је ФСХ повећан. Како фоликул сазрева, секреција естрадиола се повећава, што подстиче повећану производњу гонадотропина од стране хипофизе и појаву и ЛХ и ФСХ циклуса, тј. полни стероиди стимулишу секрецију гонадотропина.
Тренутно је утврђена структура ЛХ. Као и ТСХ, састоји се од 2 подјединице: а и б. Структура алфа подјединице ЛХ код различитих животињских врста се у великој мери поклапа, одговара структури алфа подјединице ТСХ.
Структура бета подјединице ЛХ значајно се разликује од структуре бета подјединице ТСХ, иако има четири идентична дела пептидног ланца који се састоје од 4-5 аминокиселинских остатака. Код ТСХ, они су локализовани на позицијама 27-31, 51-54, 65-68 и 78-83. Пошто бета подјединица ЛХ и ТСХ одређује специфичну биолошку активност хормона, може се претпоставити да хомологни делови у структури ЛХ и ТСХ треба да обезбеде везу бета подјединица са алфа подјединицом, а делови који се разликују по структури треба да буду одговорни за специфичност биолошке активности хормона.
Нативни ЛХ је веома стабилан на дејство протеолитичких ензима, али бета подјединица се брзо цепа химотрипсином, а а подјединица се тешко хидролизује ензимом, тј. игра заштитну улогу, спречавајући химотрипсин да приступи пептидним везама.
Што се тиче хемијске структуре ФСХ, истраживачи још увек нису добили дефинитивне резултате. Као и ЛХ, ФСХ се састоји од две подјединице, али се бета подјединица ФСХ разликује од бета подјединице ЛХ.
Пролактин
Још један хормон, пролактин (лактогени хормон), игра активну улогу у процесима репродукције. Главна физиолошка својства пролактина код сисара манифестују се у облику стимулације развоја млечних жлезда и лактације, раста лојних жлезда и унутрашњих органа. Он подстиче испољавање ефекта стероида на секундарне полне карактеристике код мужјака, стимулише секреторну активност жутог тела код мишева и пацова и учествује у регулацији метаболизма масти. Последњих година се поклања велика пажња пролактину као регулатору мајчиног понашања; таква полифункционалност се објашњава његовим еволутивним развојем. То је један од древних хормона хипофизе и налази се чак и код водоземаца. Тренутно је структура пролактина код неких врста сисара потпуно дешифрована. Међутим, донедавно су научници изражавали сумње у постојање таквог хормона код људи. Многи су веровали да његову функцију обавља хормон раста. Сада су добијени убедљиви докази о присуству пролактина код људи и његова структура је делимично дешифрована. Рецептори пролактина активно везују хормон раста и плацентни лактоген, што указује на јединствени механизам деловања сва три хормона.
Соматотропин
Хормон раста, соматотропин, има још шири спектар деловања од пролактина. Као и пролактин, производе га ацидофилне ћелије аденохипофизе. СТХ стимулише раст скелета, активира биосинтезу протеина, има ефекат мобилизације масти и подстиче повећање телесне величине. Поред тога, координира метаболичке процесе.
Учешће хормона у овом последњем потврђује чињеница наглог повећања његове секреције од стране хипофизе, на пример, када се ниво шећера у крви смањи.
Хемијска структура овог људског хормона је сада у потпуности утврђена - 191 аминокиселинска остатка. Његова примарна структура је слична структури хорионског соматомамотропина или плацентног лактогена. Ови подаци указују на значајну еволутивну блискост ова два хормона, иако показују разлике у биолошкој активности.
Потребно је нагласити високу специфичност врсте дотичног хормона - на пример, СТХ животињског порекла је неактиван код људи. То се објашњава како реакцијом између рецептора СТХ људи и животиња, тако и структуром самог хормона. Тренутно су у току студије за идентификацију активних центара у сложеној структури СТХ који показују биолошку активност. Проучавају се појединачни фрагменти молекула који показују друга својства. На пример, након хидролизе људског СТХ пепсином, изолован је пептид који се састоји од 14 аминокиселинских остатака и одговара делу молекула 31-44. Није имао ефекат раста, али је значајно премашио нативни хормон у липотропној активности. Људски хормон раста, за разлику од сличног хормона животиња, има значајну лактогену активност.
Аденохипофиза синтетише многе пептидне и протеинске супстанце које имају ефекат мобилизације масти, а тропни хормони хипофизе - АЦТХ, СТХ, ТСХ и други - имају липотропни ефекат. Последњих година посебно су истакнути бета- и γ-липотропни хормони (ЛПГ). Биолошка својства бета-ЛПГ су детаљније проучавана; поред липотропне активности, има и меланоцит-стимулишући, кортикотропин-стимулишући и хипокалцемијски ефекат, а такође производи и инсулину сличан ефекат.
Тренутно је дешифрована примарна структура овчијег ЛПГ (90 аминокиселинских остатака), липотропних хормона свиња и говеда. Овај хормон има специфичност за врсту, иако је структура централног региона бета-ЛПГ иста код различитих врста. Она одређује биолошка својства хормона. Један од фрагмената овог региона налази се у структури алфа-МСХ, бета-МСХ, АЦТХ и бета-ЛПГ. Претпоставља се да су ови хормони настали из истог прекурсора у процесу еволуције. γ-ЛПГ има слабију липотропну активност од бета-ЛПГ.
Меланоцит-стимулирајући хормон
Овај хормон, синтетисан у средњем режњу хипофизе, стимулише биосинтезу пигмента коже меланина у његовој биолошкој функцији, подстиче повећање величине и броја пигментних ћелија меланоцита у кожи водоземаца. Ове особине МСХ се користе у биолошком тестирању хормона. Постоје две врсте хормона: алфа- и бета-МСХ. Показано је да алфа-МСХ нема специфичност за врсту и има исту хемијску структуру код свих сисара. Његов молекул је пептидни ланац који се састоји од 13 аминокиселинских остатака. Бета-МСХ, напротив, има специфичност за врсту, а његова структура се разликује код различитих животиња. Код већине сисара, молекул бета-МСХ се састоји од 18 аминокиселинских остатака, а само код људи је продужен од аминокиселинских крајева за четири аминокиселинских остатака. Треба напоменути да алфа-МСХ има извесну адренокортикотропну активност, а његов ефекат на понашање животиња и људи је сада доказан.
Окситоцин и вазопресин
Вазопресин и окситоцин, који се синтетишу у хипоталамусу, акумулирају се у задњем режњу хипофизе: вазопресин у неуронима супраоптичког једра, а окситоцин у паравентрикуларном једру. Затим се преносе у хипофизу. Треба нагласити да се прекурсор хормона вазопресина синтетише прво у хипоталамусу. Истовремено се тамо производе протеини неурофизин типа 1 и 2. Први везује окситоцин, а други вазопресин. Ови комплекси мигрирају у облику неуросекреторних гранула у цитоплазми дуж аксона и доспевају до задњег режња хипофизе, где се нервна влакна завршавају у васкуларном зиду и садржај гранула улази у крв. Вазопресин и окситоцин су први хормони хипофизе са потпуно утврђеним аминокиселинским низом. По својој хемијској структури, они су нонапептиди са једним дисулфидним мостом.
Разматрани хормони производе различите биолошке ефекте: стимулишу транспорт воде и соли кроз мембране, имају вазопресорски ефекат, појачавају контракције глатких мишића материце током порођаја и повећавају секрецију млечних жлезда. Треба напоменути да вазопресин има већу антидиуретску активност од окситоцина, док овај други има јачи ефекат на материцу и млечну жлезду. Главни регулатор секреције вазопресина је потрошња воде; у бубрежним тубулама се везује за рецепторе у цитоплазматским мембранама са накнадном активацијом ензима аденилат циклазе у њима. Различити делови молекула су одговорни за везивање хормона за рецептор и за биолошки ефекат.
Хипофизна жлезда, повезана преко хипоталамуса са целим нервним системом, обједињује ендокрини систем у функционалну целину, учествујући у обезбеђивању константности унутрашњег окружења тела (хомеостаза). Унутар ендокриног система, хомеостатска регулација се спроводи на основу принципа повратне спреге између предњег режња хипофизе и „циљних“ жлезда (штитна жлезда, кора надбубрежне жлезде, гонаде). Вишак хормона који производи „циљна“ жлезда инхибира, а његов недостатак стимулише лучење и ослобађање одговарајућег тропског хормона. Хипоталамус је укључен у систем повратне спреге. Управо у њему се налазе рецепторске зоне осетљиве на хормоне „циљних“ жлезда. Специфичним везивањем за хормоне који циркулишу у крви и променом одговора у зависности од концентрације хормона, хипоталамички рецептори преносе свој ефекат на одговарајуће хипоталамички центре, који координирају рад аденохипофизе, ослобађајући хипоталамички аденохипофизотропне хормоне. Стога, хипоталамус треба сматрати неуроендокриним мозгом.